장수와 항노화
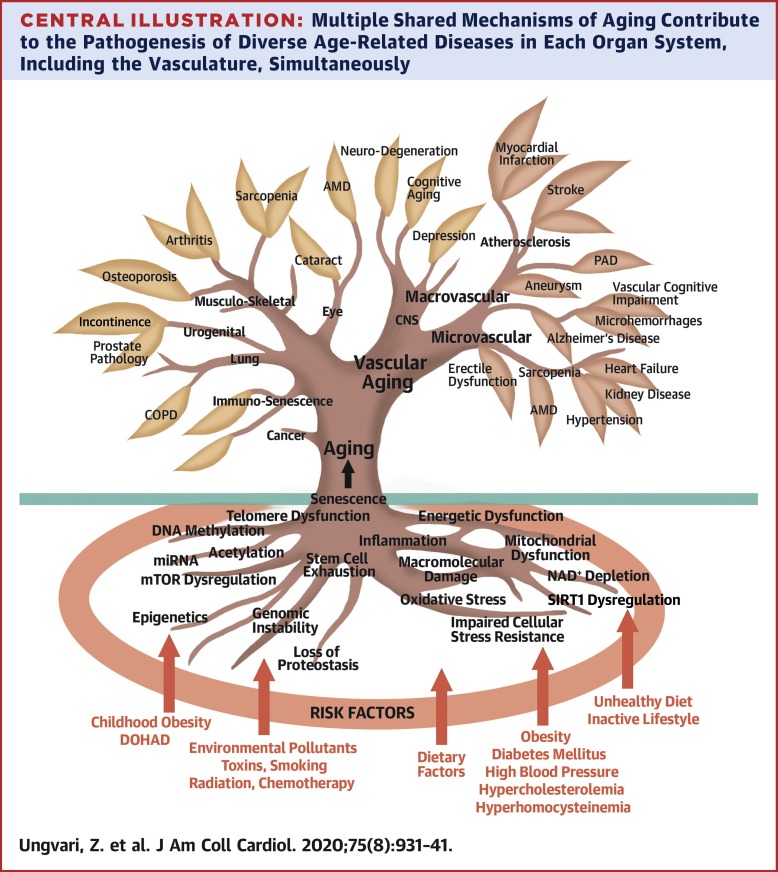
노화의 기본 메카니즘
노화의 기본 메커니즘
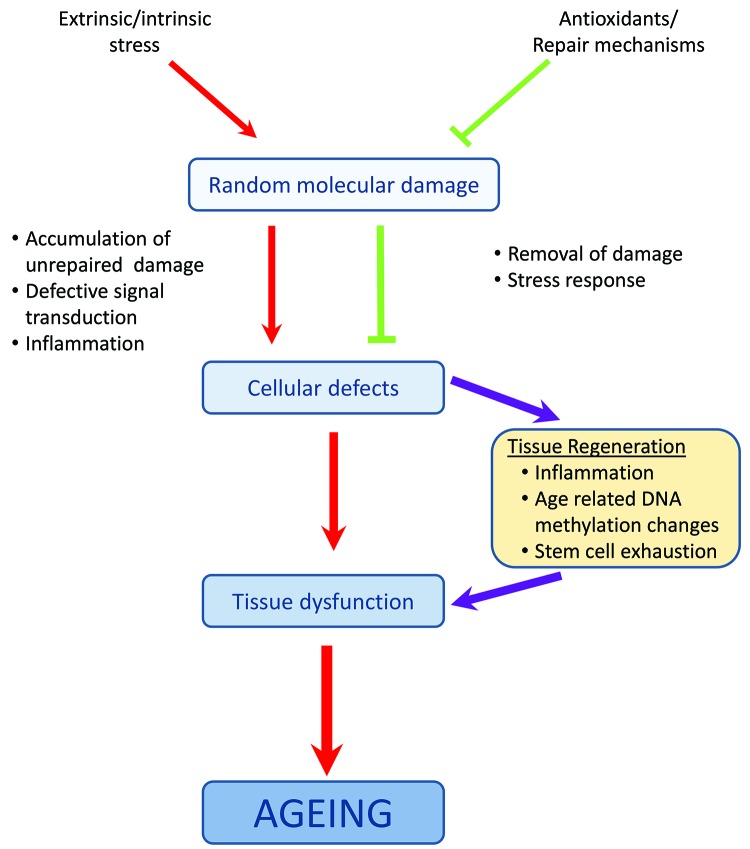
스트레스로 인한 무작위 분자 손상의 축적 속도는 항산화 시스템의 능력과 복구 시스템의 효율성에 따라 달라집니다. 이러한 시스템은 100% 효율적이지 않기 때문에 세포에는 항상 스트레스 반응의 활성화와 손상을 제거하거나 세포 분열을 방지하기 위한 메커니즘의 상향 조절로 이어지는 일부 복구되지 않은 손상이 포함되어 있습니다. 그러나 이러한 반응은 나이가 들수록 효율성이 떨어지기 때문에 손상된 구성 요소가 축적되어 세포 결함으로 이어지며, 이는 조직 기능 장애 및 노화를 유발합니다
노화의 분자 메커니즘에 대한 많은 연구는 체세포 돌연변이의 축적, 텔로미어 단축, 단백질 손상 또는 미토콘드리아 기능 장애와 같은 특정 이론에 초점을 맞춰 왔습니다. 그러나 1990년대 후반에는 개별 메커니즘이 노화 과정을 적절하게 설명할 수 없으며 이러한 다양한 메커니즘 간의 상호 작용을 고려해야 한다는 사실 이 인식되었습니다
예를 들어 손상된 미토콘드리아는 더 많은 ROS를 생성하여 결과적으로 모든 분자 구성 요소의 손상을 증가시킵니다. 이로 인해 노화에 대한 네트워크 이론이 탄생했고 복잡한 상호작용을 연구하는 과제가 노화에 대한 최초의 통합 수학적 모델 중 하나를 탄생시켰습니다. 그 이후로 많은 새로운 기술의 출현과 대량의 실험 데이터를 생성할 수 있는 능력으로 인해 분석과 해석을 돕기 위한 새로운 도구의 개발이 필요했고, 이는 시스템 생물학 접근법의 출현으로 이어졌습니다
노화의 노화의 분자 메커니즘 모델
2. 텔로미어 단축
텔로미어는 선형 염색체의 끝을 보호하는 반복적인 DNA 서열입니다. 인간 체세포에서 말단 복제 문제로 인해 각 세포 분열마다 텔로미어가 짧아지며, 텔로미어 단축은 인간 이배체 섬유아세포가 세포가 분열을 겪기 전에 제한된 횟수(헤이플릭 한계로 알려짐)만 분열할 수 있는 이유입니다.
산화 스트레스가 텔로미어 단축 속도를 최대 10배까지 증가시키는 데 기여하는 중요한 요소입니다
3. 단백질 항상성 상실
단백질의 항상성은 세포 기능에 중요하며 단백질 합성, 접힘 및 재접힘과 관련된 품질 관리 시스템에 의해 유지됩니다.
노화에서 손상된 단백질의 역할에 대한 증거는 뇌에서 나이가 들수록 산화된 단백질의 증가가 관찰된다는 것입니다.
단백질은 형태적 또는 공유결합 변화를 초래하는 산화 손상에 취약합니다. 대부분의 공유 결합 손상은 되돌릴 수 없으므로 이러한 손상이 있는 단백질은 축적과 교차 결합을 방지하기 위해 분해되어야 합니다. 구조적 손상은 노출된 소수성 표면에 결합하고 다시 접히는 것을 돕는 분자 샤페론에 의해 복구될 수 있습니다. 그러나 나이가 들면서 샤페론 시스템이 압도되어 손상되거나 잘못 접힌 단백질이 추가로 증가한다
1) 분자보호자 – 샤페론
스트레스 조건 하에서는 Hsp90에 결합하여 HSF-1을 방출하는 잘못 접힌 단백질이 증가하며, HSF-1은 삼량체화되어 핵으로 이동하여 전사적으로 활성화됩니다. 이로 인해 변성된 단백질을 다시 접는 데 도움을 줄 수 있는 분자 샤페론이 증가합니다.
만성 스트레스는 결국 분자 샤페론과 잘못 접힌 단백질 사이의 균형이 유지되지 않는 지점으로 이어졌고, 잘못 접힌 단백질은 서로 결합하여 응집체를 형성했습니다. 잘못 접힌 단백질의 수준이 높은 세포가 프로그램된 세포 사멸을 겪을 수 있는 가능성이 높아집니다
2) 단백질분해 경로
단백질 분해에는 자가포식 시스템과 프로테아솜 시스템이라는 두 가지 주요 경로가 있습니다. 두 시스템 모두 손상된 단백질을 제거할 수 있습니다.
3) 단백질 응집
연령 관련 신경퇴행성 장애의 단백질 응집에 대한 많은 모델이 개발되었습니다. 일부 모델은 응집 과정의 역학에만 초점을 맞추는 반면, 다른 모델은 단백질 분해, 샤페론, 신경염증, DDR , 산화 스트레스 및 세포사멸 경로. α-시누클레인의 응집은 PD의 핵심 요소입니다.
미토콘드리아 기능 장애는 산화 스트레스 증가, 에너지 손상 및 자가포식 억제로 이어지기 때문에, 미토콘드리아 손상과 ROS(다음 섹션 참조) 모델과 단백질 응집을 연결하면 노화와 관련된 단백질 동질성 감소의 원인에 대한 새로운 통찰력을 제공할 수 있습니다.
4. 미토콘드리아 손상 및 ROS
Harman이 노화에 대한 자유 라디칼 이론을 확장하여 세포 내 ROS 생산의 주요 부위인 미토콘드리아를 포함시킨 이후 , 증가된 ROS 수준과 기능 장애가 있는 미토콘드리아가 노화 및 연령 관련과 연관되어 있다
노화는 손상된 미토콘드리아의 축적과 관련이 있으며, 이는 미토콘드리아 회전율의 감소로 인해 발생할 수 있습니다.
5. 세포 신호의 조절 장애
음식(영양소)가 장수 스위치(장수와 관련된 많은 4가지 생물학적 경로를 조절한다..
- 인슐린 신호 전달 경로
- 두 번째는 mTOR의 기능 장애입니다
단식을 하면 mTOR가 억제되어 재활용 정리의 중요한 부분인 자가포식이 가능해집니다. 그러나 근육을 형성하려면 mTOR 활성화도 필요하며, 근육 합성을 자극하려면 류신이 포함된 실제 단백질이 필요합니다 - AMPK, 즉 아데노신 모노포스페이트 키나제입니다
결핍의 영향을 받는 효소로 낮은 에너지 상태를 감지하면 신체가 AMPK를 활성화합니다. 그러면 실제로 이러한 모든 장수 경로가 활성화되고, 이것이 바로 메트포르민이 작동하는 곳입니다 - 시르투인 ; 레스베라트롤과 같은 다양한 식물화학물질에 의해 활성화됩니다
그리고 실제로 DNA 복구를 활성화하고, NF 카파 B를 억제하고, 미토콘드리아 생물 발생을 유도하는 등의 작업을 수행합니다.
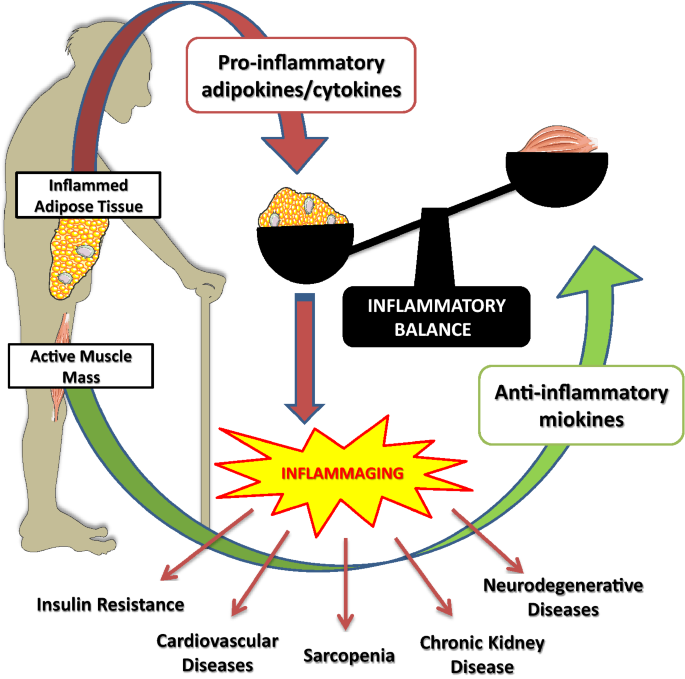
음식은 염증, 미토콘드리아 기능 장애, 미생물군집 변화, 노화 판매, 텔로미어 단축, 후성 유전적 변화, 줄기 세포 고갈, 단백질 손상, 단백질 영향, 직장 조직 장애, 단백질 당화와 같은 많은 현상, 자가포식 장애 등에 영향을 줍니다
염증이나 미토콘드리아 기능 장애에는 독소의 영향이 많습니다 .
장수와 노화도 기능의학적 관점에서 보면 그 해결책이 만성질환, 난치성질환의 기능의학적 치료법과 비슷합니다 ..

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}